生物多様性研究の意義
生物多様性の価値
周知のように生物多様性は生態系,種,種内の大きくは三つの内容をもっている。それぞれについて多くの未解決の課題がある。ここではこのうちの主に種の多様性について述べる。 種の多様性の保全はなぜ必要なのだろうか。生物種はどれくらいの速度で減少しているのだろうか。その原因は主に何であるか。それはどうすれば防ぐことができるか。以上は種の多様性に関する疑問であり,課題であり,解答である。
なぜ必要かについては互いに関連した二つの答えがあろう。あらゆる生物はその存在の基盤を生態系においている。また,生物多様性はその生態系の重要な構成 要素の一つである。したがって人間を含める生態系の保全が不可欠である以上,生物多様性の保全もまた不可欠である。既に幾多の出版物で現在の生物種の減少 速度は恐ろしく速いことが指摘されている。しかし,この地球上にどれくらいの種がいて,それが毎日,毎年どれくらいの速度で変化(減少)しているかについ ては概括の値が分かっているだけで,こと一つの地域になるとさっぱりとわかっていない。
広い意味で生物多様性の価値は生態系の諸機能に基づいている。したがって人にとっての生態系の諸機能を述べればすなわち生物多様性の価値を述べたことにな ろう。表2-1はそれをまとめたものである。人の消費的な利用価値と生産物を利用する場合を直接的な価値とし,環境維持に関するもの,動植物を通じての情 緒的な,文化的な価値などを間接的な価値としてある。有機物質の生産や無機物質・水のリサイクルなどはひろく生態系のサービスとも呼ばれる。生態系のなか での生物要素の存在は種と種群つまり群集である。したがって系の持続性などを議論する場合には生物要素としては種群を対象として考えなければならないこと になる。
|
表2-1 生物多様性の人の生活にとっての価値
|
| 直接的価値(消費的利用価値と生産的利用価値) |
| (1)太陽エネルギー獲得者としての緑色植物の保全 |
| (2)有用な遺伝形質をもった生物 |
| 間接的価値(非消費的利用価値) |
| (1)植物による水サイクルの維持(地下水の涵養,流域の保全,極端な過不足を緩和する) |
| (2)植生によるマクロ,ミクロの気候レベルでの気候の調整(気温・降水量・乱気流など) |
| (3)樹林などによる土壌の造成,侵食の防止 |
| (4)植物や土壌動物による炭素・窒素・酸素などの基礎的元素の保存と循環,および酸素と二酸化炭素とのバランスの維持 |
| (5)さまざまな汚染物質の生物による吸収と分解(微生物,植物) |
| (6)自然環境のレクリエーション的・美的・社会文化的・学術的・教育的・精神的・歴史的価値の提供など,人が自然との主に視覚的な交流を通じて得られる利益 |
| 「撹乱に抵抗する力」 |
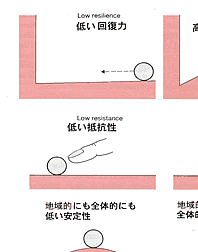
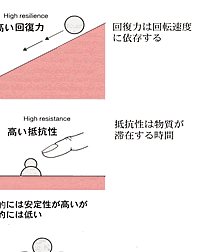
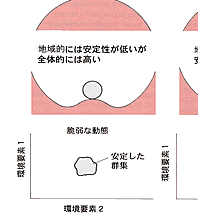
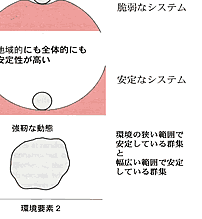
| 生態系の持続性を阻害するものは撹乱である。撹乱には自然的な力,人為的な力があるが,ここでは当然後者が問題である。ベゴンら(Begon, et al.,1986)は生態系(かれらは群集としているが,考え方としては生態系でも適応可能である)は環境改変つまり撹乱に対して回復力,抵抗力,安定性をもっているとして図2-1のように表している。 回復力は系が撹乱され,状態を変えられたときに最初の状態に戻るスピードで表現する。抵抗力は生態系に対して外力が働いたとき,その外力の影響を受けに くい力である。図2-1で表したように二階建てで地面にめり込んだように,構造が強固であれば抵抗力は大きい。安定性はその群集の存在する基盤の大きさに 関係する。図では地方的と広域的の二つの基盤を想定している。二つの基盤ともに脆弱でその上に乗った生態系は少しの外力でも変化してしまう場合,地域的に は安定性は高いが広域的には低い場合,その逆の場合,二つの基盤とも高い場合の4つの場合が考えられる。 例えば尾瀬の湿原を考えれば,回復力,抵抗力ともに弱いが,安定性は高いと言えるであろう。また,珊瑚礁生態系は回復力は高いが広域的には弱いと言えるか もしれない。ただし具体的には外力のかかり方,その強さ,生態系のサイズによってこの図のどれに当てはまるかがきまる。要は当てはめてみることによって生 態系の三つの性質を計測し,撹乱要因を排除するための具体的な方策を立てることにある。実際には対象とする生態系の要素である生物群集自体がもっている諸 性質とそれが存立している環境の状態とが群集の安定性に関係する。そこで問題は生態系を構成する種群と系の性質との関係である。その一つとして生物群集を 構成する種の数が増加すれば系は安定するか?という問題について次に述べる |
 |
 |
 |
 |
|
図2_1 生物郡集の回復力、抵抗性、安定性のモデル
(Begon et al., 1986) |
|
| 「多様性と安定性について」 |
この問題は現在でも多くの研究が行われ,かつ議論がなされている課題である。議論の出発点はC.エルトンの種類数が多くなれば生物群集の安定度は増加す る,という命題である。これはエルトンの慣習的知識(Conventional Wisdom)と呼ばれる(Elton, 1954)。この場合に群集の安定度を何で表すかということが問題であり,それを巡っての議論が今なお決着していない理由である。先のベゴンらは両者に明 確な相関がない場合とある場合の5つの事例をあげて結論はケースバイケースで異なるとしている。因みにその5つとは
つまり,種数と群集の安定性に関しては明確な結論はまだ得られていないのである。それぞれ異なった生活型をもった種群で種間関係のあり方を具体的に分析 してみることから,この問題は再出発すべきであるということである。近年は保全生物学の発展とともにこの問題は新たな脚光を浴びつつあるように思える。例 えば,Siemannら(1998)は植物の種多様性の増加は無脊椎動物の種多様性を増加させるが,部分的な草食性の多様性は,順次に,寄生者または捕食 者の多様性を保っていることを実験的に確かめ,Hubbeltら(1999)はギャップの存在が熱帯樹林の種多様性を保つのに一定の効果をもっていること を分析している。私の手元のデータベースでもNature誌,Science誌だけ見ても1999年には11,2000年9月までに17編の論文がこの問 題を論じている(Ivesら,1999:Hectorら,1999)。 |
| 「レッド・データ種のもつ意味」「絶滅危惧種の課題は生物多様性の課題でもある」 |
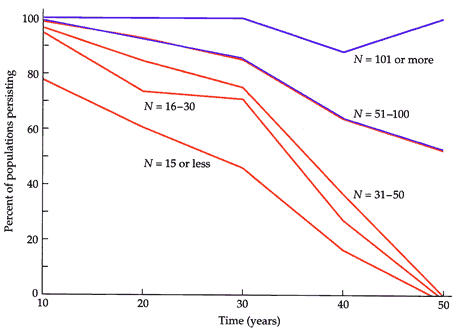
| 「国際自然保護連合(IUCN)」は2000年9月28日に「レッドリスト2000」を発表した。全世界の絶滅危惧種 は,哺乳類1030種,鳥類1083種,爬虫類296種,両生類146種,魚類752種,無脊椎動物1928種とされた。今回の調査から新基準をあてはめ た植物は,5611種に上った。絶滅の恐れがある動物が1996年版に比べ230種増えるなど,生態系破壊の進行を裏付ける結果となっており,IUCNは 「極めて深刻な事態」と警告している。 当然のことながら絶滅危惧種はこれを安全な個体数まで回復させるためのメルクマールとしての意味があるのであり,指定することそのものに意義があるわけで はない。危機にある種の救出のための生物学は保全生物学である。プリマック(Primack, 1998)は“今日では人の活動によって種の絶滅の早さは自然でのそれの1000倍にも速くなっている-それは新種の誕生とは比べものにならないほど速 い。絶滅が危惧されているような種ではわずかの個体群や時にはたった1個体群が生き残っていることもあるが,それらはその種の保全にとって核である。ある 環境条件下でのそれらの個体群の安定性が重要である”と述べ,存続可能最小個体群(minimum viable population: MVP)に基礎をおく保全の理論を展開した。ここで存続可能最小個体群とは,種の長期間の生存を保証するのに必要な個体数のことで,予測可能な未来にまで 高い確率で存続できることが保証されるような一番小さな個体群サイズのことである。最良の例は,南西アメリカでのオオツノヒツジ(Ovis canadensis)であるとされている(図2-2)。驚くことに,50頭以下の個体群は50年以内に100絶滅したが,100頭以上は現在でも存続 していることが明瞭にわかる。 プリマックはMVPに関して,個体数は多い方がよいが,小個体群が絶滅する要因としては,①遺伝的多様性の喪失と,近交弱勢や遺伝浮動を起こす,②出生 と死亡のランダム変動による人口学的変動,③捕食,競争,病気,食物供給などの変動による環境変動,④火災,洪水,噴火,台風,干魃などの不定期の自然災 害を挙げている。これに関連して,遺伝的変異性の喪失,個体群サイズの効果など個体群生態学的な,また集団遺伝学的な調査・研究の必要性を説いている。 今回の福岡県のRD種検討ではこれらの個体群的な検討は全く行っていない。また,オオツノヒツジの例のような絶滅危惧曲線を書けた例もない。これは過去の 分布が不明な種が多いことにも原因があるが,個体群の定量という点での注目度が低かったことに起因するものである。次のRD種を検討する機会には数種につ いてでもこれらの検討が行われることが望まれる。 |
 |
|
図2_2 オオツノヒツジ個体群の時間(年)と個体群の残存確率()
(Primack, 1998) |
| 「体の大きさの課題」 |
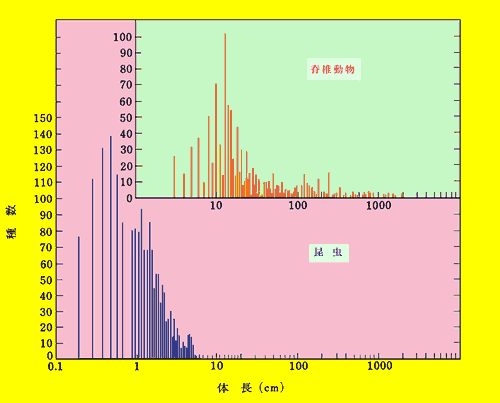
| MVPについてはその存続を保証する生息地の大きさが不可欠である。MVPの互いに関係しあっている全体はメタ個体群 と呼ばれるが,メタ個体群を維持するのに必要な面積を最小要求面積(minimum area requirement: MAR)という。最小要求面積は当然のことながら対象とする動物の大きさで異なる。このことを考えに入れることがRD種の保全には不可欠であろう。図 2-3は北米の一地域での全動物についてサイズの頻度分布を表したものである。昆虫類と脊椎動物とでは頻度分布は全く異なった分布型となる。つま り,MVPのあり方は少なくともこれらの違いを考慮に入れなければならないと言えよう。従来RD種に関する議論では無脊椎動物も脊椎動物も同じRD種に 入っているということだけで,その生活型の相違についてはほとんど議論されてこなかったが,生息地のあり方に関しては今後このような考え方は不可欠であろ う。今回,福岡県RD種を定めるに当たって重要な植物群落の存在に注目したのも以上の理由による。 |
|
(小野勇一)
|
 |
|
図2_3 北米における脊椎動物と昆虫類の体調の種類頻度分布
(Pianka, 1971) |
| (参考文献) |
| Siemann, E., D. Tilman, J. Haarstad I and M. Ritchie. 1998. Experimental tests of the dependence of arthropod diversity on plant diversity. Amer. Nat. 152:738-50. Elton, C. S. and R. S. Miller. 1954. The ecological survery of animal communities: with a practical system of classifying habitats by structural characters. J. Ecol. 42:460-496. Hubbelt, S. P., A. B. Foster, S. T. O'Brien, T.K.E. Harms, R. Condit, B. Wechster, S. J. Wright, S. Loo de Lao. 1999. Light-gap disturbances recruitment limitation, and tree diversity in a neotropical forest. Science 283:368-70. Ives, A. A., K. Gross and J. L. Klug. 1999. Stability and variability in competitive communities. Science 286:542-4. Hector, A., B. Schmid, C. Beierkuhnlein, M. C. Caldeira, M. Diemer, P. G. Dimitrakopoulos, J. A. Finn, H. Freitas, P. S. Giller, J. Good, R. Harris, P. Hogberg, K. Huss-Danell, J. Joshi, A. Jumpponen, C. Korner, P. W. Leadley, M. Loreau, A. Minns, C. P. H. Mulder, G. O'Donovan, S. J. Otway, J.S. Pereira, A. Prinz, D. J. Read, M. Scherer-Lorenzen, E.-D. Schulze, A.-S. D. Siamantziouras, E. M. Spehn, A. C. Terry, A. Y. Troumbis, F. I. Woodward, S. Yachi, and J. H. Lawton. 1999. Plant diversity and productivity experiments in European grasslands. Science, 286:1123-27. Primack, R. B. 1998. Essentials of conservation biology (2nd Ed). Sinauer Assoc. Mas. USA. Begon, M., J. L. Harper and C. R. Townsend. 1986. Ecology: individuals, populations and communities. Blackwell Scientific Publication. |